树木生长模型
1.森林生长方程
(1)Leak和Graber(1976):

(2)Solomon(1981):

(3)Beck(1974):
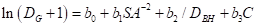
(4)Arney(1971)之后的Hegyi(1974):

(5)Belcher等人(1982):

(6)Lemmon和S出Macher(1962):

(7)Ek和Monserud(1974):

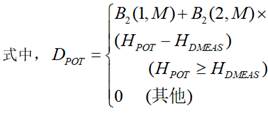

森林生长模型中的符号; 是直径的增量;
是直径的增量; 是胸高
是胸高
直径; 是基部面积;
是基部面积; 是第i高度的累积树干体积的增长;
是第i高度的累积树干体积的增长;
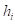 是地面上的第i高度;
是地面上的第i高度; 是回归预测的生长系数;
是回归预测的生长系数;
是5年的直径增长;S是生境指数;A是初始年龄;C是竞争指数;
 是离树梢距离为
是离树梢距离为 处的直径;
处的直径;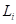 是从树梢到
是从树梢到 测量处的
测量处的
以英尺为单位的长度; 是在
是在 测量点处以年为单位的年龄;
测量点处以年为单位的年龄;
 是树冠的比率代码;
是树冠的比率代码; 和
和 是特定物种的回归系数;
是特定物种的回归系数;
 是对y的截留,被用作生长潜力中的误差修正因子;
是对y的截留,被用作生长潜力中的误差修正因子; 是对于
是对于
物种可预测的每英亩中的最大基部面积;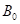 是一个把树木的实际
是一个把树木的实际
直径和平均直立直径相关联的函数;Y越来表示一棵树在5年和10
年期间的每年生长量;D是一棵树当前的胸高直径;H是树木目前的
总高度;A是树木目前的年龄;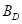 是领地周围的基本面积;
是领地周围的基本面积;
是测量直径的高度;B是净树干长度;H(I)是初始高度; 是用
是用
忍耐力调整了的竞争指数; 是相关的随机变量;S(M)是索引
是相关的随机变量;S(M)是索引
年龄中的平均优势高度; 是高度-年龄方程;
是高度-年龄方程; 是开放
是开放
生长的直径-高度方程的参数; 是用竞争调整直径增加量的参数;
是用竞争调整直径增加量的参数;
 是用竞争调整高度增加量的参数。
是用竞争调整高度增加量的参数。
2.群落动态增长方程
(1)基于合成代谢和分解代谢率的增长率(Pienaar 和Turnbull
(1973)根据Von Bertalanffy(1951))――增长的潜在速率:
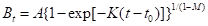
式中, 用于基本的面积、高度和体积;A是
用于基本的面积、高度和体积;A是 的渐近线的值;K,
的渐近线的值;K,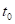 和
和
M是迭代过程产生的合适数据的参数值。
(2)没有倍增限制的增长(Phipps(1979)):
1)直径增量:

2)拥挤、遮蔽和遮蔽忍耐力:

3)土壤湿度:

式中, 是第n环增长的宽度;
是第n环增长的宽度; 是距离前一环外围的半径长度;
是距离前一环外围的半径长度;
P是在圆环从圆锥形变成抛物面形的变形点处的半径长度;H的湿度
影响的倍增乘数;G是拥挤乘数;n是被测圆环的数量;M是树干密度
的参数;t是遮蔽忍耐力参数;T是取样点的地下水深度;W是物种的最
佳地下水深度; 是比较100年时间序列的实际圆环宽度和理论圆环
是比较100年时间序列的实际圆环宽度和理论圆环
宽度比率得出的气候噪音乘数。
(3)有倍增限制的增长――最佳增长方程:
1)Botkin等人(1972):

2)Dale和Hemstrom(1984):
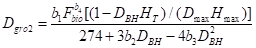
式中, 是最佳直径增长;
是最佳直径增长; 是树木的高度;
是树木的高度;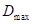 是物种的增长
是物种的增长
参数;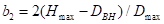 ;
; ;
;
是树叶生物量; 是参数。
是参数。
3)光对直径增长的影响(Botkin等人,1972):

式中,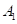 是一棵树木接收到的有效光照;
是一棵树木接收到的有效光照;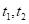 是特定物种或物种的
是特定物种或物种的
光照影响参数。
4)温度对直径增长的影响(Botkin等人,1972):

式中, 表示积温(
表示积温( )
) 是一个物种能够忍受的积温的
是一个物种能够忍受的积温的
最小值; 是一个物种能够忍受的积温的最大值。
是一个物种能够忍受的积温的最大值。
(4)水应力对直径增长的影响:
1)Solomon和Shugart(1984):

2)Pastor和Post:

3)Reed和Clark(1979):

式中, 是每个生长季节的干燥天数;
是每个生长季节的干燥天数; 是生长季节的长度;
是生长季节的长度;
 是一个物种能够忍受的土壤湿度在萎蔫点以下的生长季节的
是一个物种能够忍受的土壤湿度在萎蔫点以下的生长季节的
比例; 是负的木质部水势;
是负的木质部水势; 是物种水应力的最小值;
是物种水应力的最小值;
O是对于生长的最佳木质部水势; 是负x截留。
是负x截留。
(5)营养压力和直径增长:
1)Aber等人(1979)根据Mitchell和Chandler(1939):

2)Weinstein(1982)根据Mitchell和Chandler(1939):

式中,N是相对的氮的可用性; 和
和
 是物种组的参数;
是物种组的参数; 是如果树木生长不受营养条件的
是如果树木生长不受营养条件的
限制,可交换的营养储量将要减少的比例。
3.竞争指数的数学表达式及其作者
(1)立地密度的度量:
1)Beck(1974):

2)Botkin等人(1972):
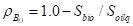
(2)影响区域的重叠:
1)Gerrard(1969):

2)Bella(1971):

(3)距离加权大小的比率:
1)Hegyi(1974):

2)Daniels(1976):

3)Moserud和Ek(1977):

(4)生长多边形(Doyle,1983):

式中, 是第j竞争者的基部面积;n是竞争者的总数;
是第j竞争者的基部面积;n是竞争者的总数; 是
是
总的立地基部面积; 是有记录的森林基部面积的最大值;A是
是有记录的森林基部面积的最大值;A是
影响区域的面积;O是重叠的面积;D是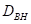 ;
; 是指数因子;
是指数因子;
 是树木之间的距离;¢是估算竞争速率的参数;
是树木之间的距离;¢是估算竞争速率的参数;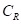 是开放增长
是开放增长
树冠半径; 是竞争者对其影响者的树冠影响的叶腋内角度。
是竞争者对其影响者的树冠影响的叶腋内角度。
参考文献
Dale V H, Doyle T W,Shugart H H: 树木生长模型的比较。
EcologicalModelling,1985,29






